Mécanismes de défense de l'organisme
L'organisme est constamment exposé à une variété de facteurs qui peuvent potentiellement l'endommager; ce sont les bactéries, les virus et certains composés chimiques. Par exemple, les bactéries pathogènes envahissant l'organisme libèrent soit des enzymes susceptibles de détruire les membranes et les organites cellulaires, soit les toxines qui modifient les fonctions normales des organes et des tissus. Les virus peuvent pénétrer dans les cellules et utiliser le matériel cellulaire afin de reproduire d'autres particules virales. Ils peuvent tuer les cellules en les privant des composés essentiels à leur croissance ou en les induisant à produire des substances toxiques.
Heureusement, l'organisme résiste à l'envahissement de plusieurs micro-organismes et substances chimiques qui endommagent les tissus. Cette propriété s'appelle l'immunité. Elle est d'abord assurée par des mécanismes de défense non spécifiques. Ces mécanismes ne nécessitent pas une exposition préalable à une substance antigénique étrangère particulière pour s'en défendre. Ils comprennent les barrières formées par les surfaces (fig.1) et les membranes cellulaires (fig.2) ainsi que les réponses inflammatoires (voir plus loin) qui se développent à la suite d'une lésion tissulaire.
Figure 1: Coupe de muqueuse des voies respiratoires. On remarque l'épaisseur du tissu, constitué de plusieurs couches de cellules, dont certaines secrétent du mucus protecteur (en bleu en bas).
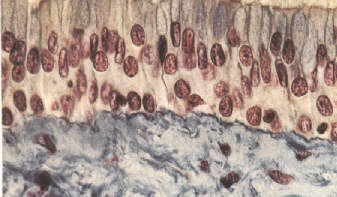
Figure 2: Illustration d'une membrane de cellule animale, dont l'espace extracellulaire est en haut, le cytoplasme en bas.
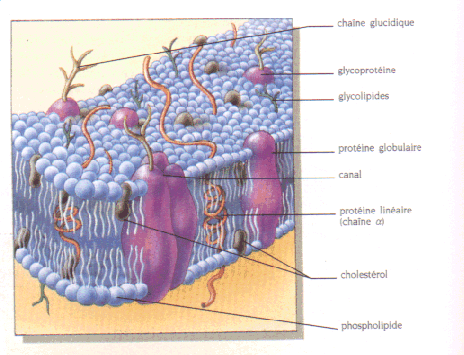
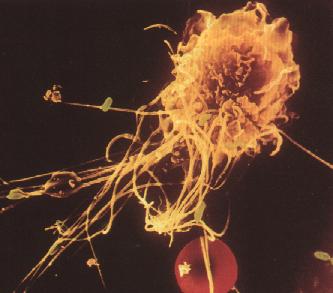
Figure 3: Macrophage phagocytant des bactéries (en vert) et un globule rouge (en rouge):
L'immunité est aussi assurée par des réponses spécifiques qui dépendent de l'exposition à une substance étrangère antigénique, de la reconnaissance de cette substance lors d'une exposition ultérieure et, enfin, de la réaction particulière à cette substance. Ces réponses immunes spécifiques sont assurées par des lymphocytes spécialisés et par d'autres cellules du système immunologique (fig.3).
La figure 5 illustre la coopération qu'il existe entre les principaux acteurs de la réponse immunitaire. Un agent infectieux (microbe) est reconnu par les macrophages et phagocyté. Découpé en fragments antigéniques à l'intérieur du macrophage, ceux-ci sont externalisés de façon à se retrouver sur la membrane du macrophage. Ils sont alors présentés aux autres cellules du système immunitaire (lymphocytes T et B), qui vont ainsi "apprendre" à reconnaître l'agent infectieux avant même de l'avoir rencontré.
Deux voies d'activation des lymphocytes existent selon la nature antigénique du microbe:
-
-
activation directe des lymphocytes B; l'antigène constituant le microbe
est dit thymo-indépendant,
- activation préalable des lymphocytes T puis, par l'intermédiaire de ces derniers, des lymphocytes B; l'antigène est dit thymo-dépendant.
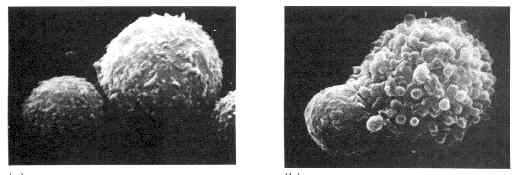
Figure 4: un lymphocyte T effecteur (activé) s'attache à une cellule reconnue comme étrangère à l'organisme (image de gauche). Celle-ci est détruite alors que de profonds replis apparaissent à sa surface, témoignant de l'action du lymphocyte (image de droite). Le lymphocyte est la petite cellule du bas à gauche des images.
L'activation des lymphocytes B les transforme en plasmocytes secréteurs d'anticorps. Grace à cette coopération entre cellules immunitaires, les anticorps produits seront spécifiques du microbe.

Quoi? Qu'est-ce que c'est que ça? Voila qu'on parle du thymus avec des antigènes thymo-duelque chose!
C'est normal puisque les lymphocytes T (T comme thymus) sont fabriqués dans cet organe. S'ils sont impliqués dans la réponse immunitaire, on dit qu'elle est thymo-dépendante, car si on ôte chirurgicalement le thymus ... il n'y a plus de réponse immunitaire possible (hormis celle des phagocytes).
Figure 5: Coopération entre cellules du système immunitaire.

SURFACES CORPORELLES
La peau et les muqueuses forment les premières barrières contre l'invasion de l'organisme par tout facteur potentiellement dommageable. Une peau sans lésion prévient l'entrée de micro-organismes et de plusieurs produits chimiques. Les glandes sudoripares et sébacées sécrètent des substances qui préviennent aussi la pénétration de plusieurs types de bactéries et la partie externe de l'épiderme contient une protéine résistante et insoluble dans l'eau (appelée kératine) en quantité suffisante pour assurer une résistance contre les acides, les bases faibles et les enzymes protéolytiques.
Les glandes muqueuses qui tapissent les cavités ouvrant sur l'extérieur, telles que les voies digestive, respiratoire (fig.1), urinaire et celle du système reproducteur, sécrètent du mucus qui piège les petites particules: celles-ci peuvent ensuite être rejetées à l'extérieur ou être éliminées par phagocytose. La muqueuse du système respiratoire supérieur est aussi tapissée de cellules ciliées qui transportent le mucus et les particules piégées vers l'extérieur. Les larmes qui baignent les muqueuses de l'oeil contiennent une enzyme, le lysozyme, possédant des propriétés antimicrobiennes; l'acidité du liquide gastrique peut détruire plusieurs bactéries et toxines bactériennes.
Même si ces barrières de surface sont très efficaces, il arrive parfois que des agents ou des organismes pénètrent dans l'organisme. Pour tenter de neutraliser ou de détruire ces agents exogènes, il est donc nécessaire que d'autres mécanismes de défense puissent entrer en jeu.
Exemple d'organisation cellulaire
pour constituer un tissu de
épidermique.
INFLAMMATION
L'inflammation est une réponse physiologique aiguë non spécifique de l'organisme à un dommage tissulaire causé par différents facteurs tels que les agents chimiques, la chaleur, les traumatismes ou l'invasion bactérienne. Même s'il existe des différences entre les agents causals, la localisation des lésions et l'état de l'organisme, la réponse inflammatoire aiguë non spécifique est fondamentalement semblable dans presque tous les cas. Ces mécanismes inflammatoires fondamentaux produisent des changements dans le débit sanguin et dans la perméabilité vasculaire et ils dirigent les leucocytes, des vaisseaux sanguins vers l'intérieur des tissus. La réponse inflammatoire aiguë non spécifique permet l'entrée, dans les zones envahies ou endommagées, de protéines plasmatiques et de phagocytes qui inactivent ou détruisent les envahisseurs, qui éliminent les débris et qui entreprennent les premières phases de la réparation des tissus. Les étapes de cette réponse supposent la participation de plusieurs substances chimiques qui sont libérées dans le tissu endommagé. Voir figure.
Les principaux symptômes de l'inflammation
Les principaux symptômes locaux de l'inflammation sont la rougeur, la chaleur, l'oedème et la douleur. La rougeur et la chaleur (augmentation de température) sont causées par la dilatation des vaisseaux sanguins et par l'augmen-tation du débit sanguin à l'endroit lésé. L'oedème est produit par l'augmentation de la perméabilité vasculaire et par le mouvement du liquide et des substances solubles, de la circulation vers les tissus. La douleur est attribuée à l'augmentation de la pression exercée sur les terminaisons nerveuses sensitives des tissus oedémateux, aux effets produits par les kinines et, peut-être aussi, à l'action d'autres médiateurs chimiques de la réponse inflammatoire sur les terminaisons nerveuses afférentes.
Symptômes systémiques
La fièvre est un symptôme systémique commun à tout processus inflammatoire, surtout aux réponses inflammatoires associées à l'envahissement de la circulation sanguine par des micro-organismes. Les neutrophiles (sous-type de globules blancs) et, peut-être, d'autres cellules associées à la réponse inflammatoire sécrètent des pyrogènes endogènes (substances provoquant l'élévation de température) qui influencent les mécanismes de régulation de la température (hausse de la température). Même si une forte fièvre peut être nuisible à l'organisme, la phagocytose est accélérée par les hausses de température. La température systémique, aussi bien que la température locale, peut augmen-ter cette activité. En plus de déclencher la fièvre, les pyrogènes endogènes permettent au fer et aux autres oligo-éléments présents dans la circulation plasmatique de passer au foie. Ce mécanisme peut retarder la prolifération des bactéries qui ont souvent besoin d'une forte concentration de fer pour se multiplier.
L'inflammation, surtout en réponse à plusieurs types d'envahisseurs bactériens, s'accompagne souvent d'une production et d'une libération accrues de leucocytes. Dans ces circonstances, le nombre de leucocytes de la circulation périphérique peut augmenter de 7000 cellules/mm3 à 25000 cellules/mm3 ou plus. De plus, au cours de la plupart des réactions inflammatoires non spécifi-ques, cette augmentation est principalement due à une croissance absolue ou relative des neutrophiles en circulation. Certains produits bactériens stimulent les monocytes et les macrophages à fabriquer une substance, la granulopoïétine, qui favorise, d'une part, la formation des granulocytes et surtout les neutrophiles par la moelle osseuse et, d'autre part, leur libération dans les tissus. Au contraire, les infections causées par les virus ou les protozoaires sont souvent associées à une diminution du nombre de leucocytes dans la circulation périphérique.
INTERFERON
L'interféron est une protéine qui joue un certain rôle actif dans la protection de l'organisme surtout au début d'une invasion virale. Les mécanismes de réponses immunes spécifiques prennent progressivement la relève. En général, lorsqu'un virus envahit l'organisme, il pénètre la cellule et produit d'autres particules virales en utilisant les constituants cellulaires pour synthétiser des composants, dont les acides nucléiques et les protéines. Les nouveaux virus sont relâchés pour réinfecter d'autres cellules saines et le processus se répète. Les virus et d'autres substances favorisent aussi la production d'interféron. Parmi les inducteurs d'interféron, l'ARN à deux brins paraît être le plus efficace. Il semble que l'ARN à deux brins puisse stimuler l'ADN cellulaire à produire de l'ARN messager qui, à son tour, contrôle la production d'interféron par la cellule. L'interféron libéré se lie à des récepteurs situés sur la membrane cellulaire de certaines autres cellules. Cette union déclenche la synthèse de protéines enzymatiques intracellulaire afin d'empêcher la reproduction virale, en décomposant l'ARN messager et en inhibant aussi la syn-thèse de protéines virales. Ces protéines enzymatiques nouvellement synthé-tisées demeurent inactivées jusqu'à ce que la cellule soit infectée par un virus ou encore exposée à de l'ARN à deux brins. Ce prérequis de l'activation sert à protéger les acides nucléiques et les mécanismes de synthèse protéinique de la cellule, en l'absence d'infection virale.
On croit que l'interféron peut aussi inhiber l'activité de certains virus par d'autres mécanismes. Même Si l'interféron ne paraît pas pouvoir intervenir dans la reproduction de certains virus, il semble pouvoir empêcher les virus synthétisés de quitter la cellule ou, si ceux-ci quittent la cellule, ils sont alors incapables d'infecter les autres cellules.
Toutes les cellules peuvent fabriquer de l'interféron lorsqu'elles sont adéquatement stimulées. Il existe au moins trois types différents d'interféron. Le premier et principal type est synthétisé par les leucocytes; le second est produit par les fibroblastes. Un troisième type, appelé interféron T ou interféron immun, est synthétisé par les lymphocytes T associés aux réponses immunes spécifiques.
L'interféron jouerait aussi un râle dans la protection de l'organisme contre certaines formes de cancer. Même si certains virus ont des propriétés oncogènes, l'interféron semble pouvoir agir contre une tumeur causée par des agents non viraux tels que les radiations ou les agents chimiques. L'interféron est considéré comme un agent versatile dont les effets sur les cellules dépas-sent de beaucoup l'inhibition de l'activité virale; par exemple, l'interféron ralentit la division cellulaire et inhibe la prolifération de cellules normales ou anormales. Il mobilise et augmente les propriétés destructives des macrophages et stimule l'activité d'un groupe de lymphocytes spécialisés appelés cellules NK (Natural Killer). Les macrophages et les cellules NK peuvent s'attaquer aux cellules tumorales. Ces deux types de cellules sont donc importants pour détecter et pour éliminer les cellules anormales qui pourraient être cancéreuses, avant que celles-ci puis-sent se multiplier et causer un cancer cliniquement détectable. On croit même que l'apparition de cellules cancéreuses serait due à l'inefficacité de ces mécanismes de surveillance immunologique. L'interféron immun paraît être un agent antitumoral plus puissant que les autres types d'interférons produits par les leucocytes ou par les fibroblastes.
REPONSES IMMUNES SPECIFIQUES
Pour être déclenchées et efficaces, les réponses immunes spécifiques requièrent généralement une préexposition à un agent étranger. Les réponses immunes spécifiques ne sont cependant pas toujours bénéfiques puisqu'elles sont parfois responsables d'allergies ou d'autres réactions indésirables.
Traditionnellement, on reconnait comme je l'ai dit précédemment deux aspects principaux aux réponses immunes spécifiques: le premier, appelé immunité humorale, est assuré par des protéines spécialisées, les anticorps. Les anticorps sont produits par les plasmocytes, qui proviennent de la lignée des lymphocytes B. Le second aspect, appelé immunité à médiation cellulaire, est assuré par certaines populations de cellules lymphoïdes, les lymphocytes T.
Antigènes
Un antigène est une substance qui peut déclencher une réponse immune spécifique lorsqu'il pénètre dans l'organisme. Les substances antigéniques ont généralement des masses moléculaires variant entre 3000 et 10000 ou plus; ce sont des protéines, des polysaccharides, des acides nucléiques ou des complexes lipidiques. Des substances de faibles masses moléculaires, appelées haptènes, peuvent déclencher une réponse immune spécifique lorsqu'elles se combinent à des molécules plus grosses, comme les protéines.
Lymphocytes B
Les lymphocytes B sont génétiquement programmés pour se différencier en plasmocytes producteurs d'anticorps associés à l'immunité humorale. Comme tous les lymphocytes, les lymphocytes B proviennent de cellules-souches précurseurs localisées dans la moelle osseuse rouge. Les cellules qui se différencient en lymphocytes B prolifèrent et se développent dans la moelle osseuse (ou dans le foie et, peut-être, dans la rate au cours de la vie foetale). Au sortir de la moelle osseuse, les lymphocytes B circulent dans le sang, les tissus et la lymphe. En tout temps, un grand nombre d'entre eux sont situés dans les ganglions lymphatiques, la rate et les autres tissus lymphoïdes.
Lymphocytes T
Les lymphocytes T comprennent un groupe de lymphocytes hétérogènes qui incluent les cellules associées aux réponses immunes à médiation cellulaire. Les cellules-souches de la moelle osseuse rouge, destinées à donner naissance aux lymphocytes T, atteignent le thymus où elles prolifèrent et se différencient. On croit que des hormones thymiques, la thymosine,
peuvent influencer les lymphocytes T qui ont déjà quitté le thymus. Tout comme les lymphocytes B, les lymphocytes T circulent dans le sang, les tissus et la lymphe; en tout temps, un grand nombre d'entre eux sont localisés dans les ganglions lymphatiques, la rate ou les autres tissus lymphoïdes.
Anticorps
Les anticorps sont des protéines spécialisées produites en réponse à une stimulation antigénique. L'anticorps peut se combiner à l'antigène spécifique qui a stimulé sa production pour former un complexe antigène-anticorps. Les anticorps appartiennent à une famille de protéines appelées globulines. On connaît cinq classes d'immunoglobulines. Les immunoglobulines G et M (IgG et IgM) sont associées à l'immunité spécifique antibactérienne et virale. Les immunoglobulines de la classe E (IgE) participent à certaines réponses allergiques. Les immunoglobulines de la classe A (IgA) proviennent des tissus lymphoïdes disséminés dans les tractus gastro- intestinal, urogénital et respiratoire; elles servent surtout à protéger ces régions. Les immunoglobulines de la classe D (IgD) sont présentes à la surface de plusieurs lymphocytes T et on croit qu'elles servent à déclencher les réponses immunes humorales.
Les anticorps sont chimiquement constitués de deux chaînes polypeptidiques "lourdes" et de deux chaînes "légères" une extrémité de chaque chaîne se trouve une région variable ou la séquence d'acides aminés diffère pour chaque type d'anticorps synthétisé. La configuration particulière des régions variables confère à l'anticorps la capacité à se lier d'une façon spécifique à l'antigène, à la manière d'une clé dans une serrure comme dans le cas de l'interrelation entre l'enzyme et son substrat. L'autre partie des chaînes polypeptidiques est appelée région constante. Les régions constantes renferment des sites permettant à l'anticorps de se fixer aux cellules ou aux molé-cules; elles agissent comme des sites effecteurs de l'activité anticorps. Les régions constantes des chaînes polypeptidiques lourdes diffèrent d'une classe d'anticorps à l'autre; ces régions permettent de distinguer les différentes classes d'anticorps.
Les anticorps inactivent et détruisent les substances étrangères de diverses façons. Ils peuvent s'unir aux toxines bactériennes ou aux enzymes étrangères nocives et les inactiver en inhibant leurs interactions avec les récepteurs cellulaires ou les substrats. Ils peuvent également se lier à la surface des virus pour empêcher ceux-ci de s'attacher à la membrane cellulaire et de pénétrer à l'intérieur de la cellule.
Une des fonctions principales des anticorps est d'accentuer la réponse inflammatoire de base. Les complexes antigènes-anticorps, particulièrement ceux formés par les anticorps de type IgG ou IgM, sont de puissants activateurs du système du complément. Comme nous l'avons déjà mentionné, les protéines activées du complément peuvent intervenir à plusieurs niveaux de la réponse inflammatoire aiguë non spécifique; ils peuvent également attaquer et tuer les micro-organismes envahissants. Les anticorps, surtout ceux du type IgC, agissent également comme opsonines pour favori-ser la phagocytose, en s'attachant aux substances étrangères; ils servent alors de structures auxquelles les cellules phagocytaires se lient.
Immunité humorale
L'immunité humorale suppose la production et la libération, dans le sang et dans la lymphe, des anticorps contre les différents antigènes reconnus comme étrangers par l'organisme. Les réponses immunes humorales sont impor-tantes parce qu'elles assurent la résistance immune spécifique contre les bactéries, les toxines bactériennes et la phase extra-cellulaire des infections virales.
Le système humoral réagit avec les antigènes de la façon suivante: lorsqu'un antigène parvient aux tissus lymphoïdes, comme les ganglions lymphatiques ou la rate, certains lymphocytes B sont stimulés et entreprennent une série de divisions cellulaires. La plupart des cellules lymphoïdes B stimulées se différencient en plasmocytes producteurs d'anticorps qui seront relâchés ensuite dans le sang et dans la lymphe. Les anticorps s'unissent aux antigènes spécifiques qui ont stimulé leur production. Apparaissent alors des réactions semblables à celles décrites dans la section précédente.
Certains lymphocytes B stimulés ne se différencient pas en plasmocytes et deviennent des cellules mémoire. Ces cellules conservent "en mémoire" l'exposition à l'antigène. Le mécanisme conduisant à la réponse humorale est relativement lent après une première exposition à l'antigène étranger et, par conséquent, il s'écoule habituellement plusieurs jours avant que la concentra-tion d'anticorps ne devienne substantielle. La mémoire immunologique sert à induire, plus rapidement une quantité supérieure d'anticorps lors d'expositions subséquentes au même antigène.
Selon une théorie sur la production des anticorps chaque type d'antigène stimule des populations différentes, ou clones, de lymphocytes B.
Chaque clone peut éventuellement produire un type d'anticorps spécifiques. Les anticorps spécifiques synthétisés par un certain clone de lymphocytes B sont exposés à la surface de ces cellules. Quand un antigène pénètre dans l'organisme, il s'unit à un des anticorps spécifiques préformés et localisés sur certains lymphocytes B. Ce phénomène stimule les lymphocytes B à se diviser et à se différencier en plasmocytes producteurs d'anticorps. Cette théorie soutient qu'il y a suffisamment de clones de lymphocytes B pour assurer une production d'anticorps spécifiques contre les millions d'antigènes auxquels une personne sera exposée au cours de sa vie.
En fait, la stimulation des lymphocytes B et la production d'anticorps nécessitent habituellement plus qu'une seule exposition à un antigène pour que les lymphocytes B commencent à produire des anticorps. La plupart du temps, les macrophages et d'autres groupes de lymphocytes T (T-Helpers) sont aussi associés à ce processus. Certains chercheurs pensent que les macrophages digèrent et présentent l'antigène aux lymphocytes T auxiliaires qui à leur tour, agissent sur les lymphocytes B pour accélérer le processus (Figure).
Immunité à médiation cellulaire
Les réponses immunes à médiation cellulaire dépendent des lymphocytes T, plus particulièrement d'un groupe de lymphocytes T appelés lymphocytes T effecteur. Ces réponses sont surtout efficaces contre les cellules qui possèdent sur leurs surfaces des antigènes que l'organisme reconnaît comme étrangers. Par exemple la plupart des cellules infectées par un virus présentent des antigènes viraux à leurs surfaces et les lymphocytes T effecteurs, produits par le système immun à médiation cellulaire en réponse à des antigènes étrangers, peuvent s'y attacher et détruire ces cellules infectées. Les réponses immunes à médiation cellulaire sont donc importantes dans la résistance immunologique spécifique, au cours de la phase intracellulaire des infections virales. Ces réponses sont également importantes pour assurer une résistance immune spécifique contre les infections causées par des parasites, des bactéries intra-cellulaires et des champignons microscopiques. Elles jouent également un râle primordial dans le rejet des greffes d'organes.
Le système immun à médiation cellulaire répond à la stimulation antigénique de la façon suivante. Tout comme pour les lymphocytes B associés à l'immunité humorale, il existe plusieurs types de lymphocytes T possédant des récepteurs de surface spécifiques pour les antigènes. Lorsqu'un antigène s'unit aux récepteurs d'un lymphocyte T particulier (les macro-phages et peut-être les lymphocytes T auxiliaires pourraient être en cause dans ce processus), les lymphocytes T se sensibilisent, entreprennent une série de divisions cellulaires et se différencient. Certains lymphocytes T provenant de cette différenciation servent à conserver en mémoire cette réponse, alors que d'autres, tels les lymphocytes T effecteurs, jouent un rôle immédiat dans la réponse immune à médiation cellulaire. Au cours de cette réponse, les lymphocytes T effecteurs quittent les tissus lymphoïdes et circulent partout dans l'organisme. Quand ils rencontrent des cellules portant le type d'antigène qui a initié leur production, ils s'unissent à elles. Ces lymphocytes T effecteurs peuvent alors détruire les cellules qui ont ces antigènes à leurs surfaces. D'autres libèrent plutôt des substances chimiques appelées lymphokines. Les lymphokines accentuent la réponse inflammatoire initiale et, par conséquent, la phagocytose. Certaines ont des propriétés de chimiotactisme, ce qui leur permet d'attirer dans la région les neutrophiles et, plus particulièrement, les monocytes qui se transforment en macrophages. D'autres lym-phokines maintiennent les macrophages sur le site de l'inflammation et aug-mentent l'activité phagocytaire; ces cellules sont appelées macrophages activés. Certaines lymphokines peuvent aussi agir comme cytotoxines et tuer directement les cellules.
Rejet de greffon
Le système d'immunité à médiation cellulaire joue un rôle important dans le rejet des tissus solides transplantés. Des antigènes, appelés antigènes d'histocompatibilité, sont présents à la surface de toutes les cellules nucléées de l'organisme. Tous les individus, à l'exception des jumeaux identiques, possèdent différents antigènes d'histocompatibilité. Lorsqu'un greffon provenant d'un individu donné (donneur) est transplanté à un autre individu (receveur) non jumeau identique, les antigènes d'histocompatibilité situés à la surface des cellules du donneur du greffon sont reconnus comme antigènes étrangers, de sorte que les lymphocytes T du receveur se sensibilisent contre ces antigènes. Ce mécanisme induit la formation de lymphocytes T effecteurs qui s'attacheront sur les antigènes présents sur les cellules du greffon et les détruiront. Lors d'une transplantation d'organe, on tentera de maintenir en échec cette immunité à médiation cellulaire assurée par les lymphocytes T effecteurs, à l'aide de divers médicaments immunosuppresseurs, comme les sérums anti-lymphocytaires, ou par l'irradiation. Dans plusieurs cas, la fonction des lymphocytes B sera également supprimée par ces traitements, de sorte que les sujets deviendront plus susceptibles aux infections.
Immunité active
La résistance à l'infection, résultant d'une activation de la réponse immune spécifique d'un individu contre un antigène donné, est appelée immunité active. Le plus souvent, l'immunité active est acquise au cours d'une infection. Elle peut également être acquise à la suite d'une vaccination, procédé qui consiste à injecter une petite quantité d'une substance antigénique à une personne. Cette substance peut être constituée de bactéries, de toxines, etc. La substance est habituellement prétraitée par séchage, par lumière ultraviolette ou par d'autres moyens, de façon à ce qu'elle ne puisse induire la maladie, tout en conservant son pouvoir antigénique pour stimuler la réponse immune spécifique.
L'activation des réponses immunes spécifiques d'un individu par l'infection ou par la vaccination permet de maintenir en alerte les anticorps et les lymphocytes T effecteurs, pour résister aux agents qui pourraient éventuellement causer la maladie. De plus, si une personne, déjà infectée ou vaccinée, est réexposée au même agent causal de la maladie, la mémoire relevant de l'immunité humorale et de l'immunité à médiation cellulaire lui permettra de répondre rapidement. Il convient de mentionner, cependant, que certains micro-organismes sont impuissants à activer le système de la mémoire immune et que la réponse à chaque exposition de l'un ou l'autre de ces micro-organismes est toujours une réponse primaire, lente comme lors d'une première exposition.
Immunité passive
L'immunité passive est en quelque sorte une immunité "empruntée". Par exemple, les anticorps dirigés contre un agent pathogène (fabriqués par un être humain et souvent même par un animal) peuvent être injectés à un individu. Malheureusement, l'immunité passive est souvent de courte durée et ne persiste que durant quelques semaines. De plus, dans l'immunité passive, les mécanismes immunologiques spécifiques du receveur ne sont pas stimulés à produire des anticorps ou des lymphocytes T effecteurs contre l'agent causal. Il existe, par exemple, au cours de la grossesse, un mécanisme d'immunité passive. Des anticorps de type IgG formés par les lymphocytes de la mère passent à travers le placenta pour atteindre le foetus. Ces anticorps protègent l'enfant jusqu'après la naissance, tant que le système immunologique de l'enfant ne peut produire ses propres anticorps. L'immunité passive peut également être acquise par l'injection d'anticorps préfabriqués. Par exemple des anticorps contre le venin d'une espèce de serpent peuvent être produits et extraits du plasma du cheval; ils peuvent ensuite être injectés a une personne qui a été mordue par ce type de serpent. L'immunité passive lui procure alors une protection immédiate contre le venin. Ce traitement n'est cependant pas sans danger: les anticorps injectés peuvent eux-mêmes s'avérer antigéniques pour le système immunologique du receveur, surtout Si ces anticorps ne sont pas d'origine humaine. Ainsi, l'injection d'anticorps peut par elle-même, sti-muler le receveur à produire des anticorps contre les premiers anticorps. Ce phénomène peut être responsable de réponses allergiques dangereuses.
Hypersensibilité
Les réactions d'hypersensibilité sont des réponses immunes qui peuvent être dommageables à l'organisme. L'immunité humorale aussi bien que l'immunité à média-tion cellulaire jouent un rôle dans ces réponses immunes.
Rôle des facteurs du complément dans l'hypersensibilité
Dans certains types d'hypersensibilité tels que l'hypersensibilité à complexe immun et l'hypersensibilité cytotoxique, les facteurs du complément peuvent induire des lésions tissulaires ou cellulaires au cours de la réponse immune humorale à un antigène. Par exemple les complexes immuns (complexes antigènes-anticorps) peuvent se déposer dans l'endothélium des vaisseaux sanguins ou ils activent le système du complément et initient une réponse inflammatoire qui endommagera le vaisseau sanguin. Le dépôt de complexes immuns dans le rein produit aussi une réaction inflammatoire connue sous le nom de glomérulo-néphrite.
Rôle des lymphocytes T effecteurs dans l'hypersensibilité
Au cours de l'hypersensibilité à médiation cellulaire (hypersensibilité retardée), l'activité des lymphocytes T effecteurs peut induire des lésions tissu-laires. Par exemple la toxine de l'herbe à puce se comporte comme un "haptène" en se liant aux protéines ou aux membranes des cellules de la peau pour sensibiliser l'organisme. En conséquence, l'exposition à la toxine de l'herbe à puce peut induire une production de lymphocytes T effecteurs contre la toxine. Lorsque l'individu sensibilisé sera réexposé à la toxine, les lymphocytes T effecteurs et les lymphokines libérées produiront une réaction inflamma-toire au site de l'exposition. Cette réaction est dommageable pour le tissu et elle s'accompagne de démangeaisons d'oedème et de vésicules. Les réactions d'hypersensibilité associées aux lymphocytes T effecteurs atteignent généra-lement leur intensité maximale entre un à trois jours.
Rôle des anticorps de type IgE dans l'hypersensibilité
Les anticorps de type IgE participent à un type d'hypersensibilité (dite anaphylactique) qui est la cause de plusieurs réactions allergiques communément rencontrées. Lorsqu'un individu est susceptible de réagir à un antigène particulier qu'il s'agisse de poussière, de pollen ou d'un allergène alimentaire on l'expose à cet antigène afin de stimuler la production d'anticorps IgE par les plasmocytes de son organisme. Ces anticorps se lient aux mastocytes et aux basophiles. Lorsqu'un individu ainsi sensibilisé est réexposé à l'antigène, ce dernier s'attache à l'anticorps présent sur la surface des mastocytes et des basophiles. Cette réaction provoque la libération d'histamine et d'autres médiateurs chimiques qui induisent une réponse inflammatoire. Il s'ensuit une vaso-dilatation, une augmentation de la perméabilité vasculaire, une augmentation de la sécrétion de mucus et une contraction des muscles lisses des bronches et des bronchioles pulmonaires.
La réaction allergique anaphylactique est souvent localisée en un endroit particulier de l'organisme. Par exemple Si l'exposition à un antigène particulier se produit dans les voies nasales, la réaction déclenchera des éternue-ments, de la rhinorrhée de la congestion et d'autres symptômes du rhume des foins dus à l'irritation, l'hypersécrétion de mucus, l'augmentation du débit sanguin et l'exsudation de protéines dans la muqueuse nasale. Occasionnellement (surtout en réponse à l'injection d'un antigène) une réaction allergique anaphylactique peut se produire; cette réaction "plus systémique que localisée" s'accompagne d'une hypotension sévère et d'une constriction rapide des bronches et des bronchioles. Cette réaction est appelée choc anaphylactique. Elle peut être fatale sans le recours à un traitement rapide (l'injection d'adrénaline par exemple). Certains individus sensibilisés développent un choc anaphylactique en réponse à la substance antigénique d'une seule piqûre d'insecte ou en réponse à certains médicaments comme la pénicilline.
L'administration d'antihistaminiques produit parfois un soulagement temporaire des réactions allergiques anaphylactiques mais ce soulagement est souvent incomplet parce que des médiateurs chimiques autres que l'histamine sont également relâchés dans le milieu lors de l'éclatement des mastocytes et des basophiles. Lorsqu'on réussit à identifier l'antigène sensibilisant respon-sable une thérapie de désensibilisation peut être entreprise. La désensibilisa-tion s'effectue par l'injection de petites quantités croissantes de l'antigène sensibilisant. Certains chercheurs pensent que ce traitement induit la production d'immunoglobuline IgG plutôt que celle d'immunoglobuline IgE contre l'antigène. Selon cette hypothèse, lorsque la personne sensibilisée rencontre de nouveau l'antigène particulier ce sont les anticorps IgG qui s'attacheront à l'antigène l'empêchant ainsi de se lier aux anticorps IgE à la surface des mastocytes et des basophiles. On pense également que la désensibilisation peut stimuler la production de lymphocytes T suppresseurs; ceux-ci suppriment la synthèse d'anticorps IgE dirigés contre l'antigène. Dans un cas ou l'autre l'antigène ne s'attache plus aux anticorps IgE liés à la surface des mastocytes et des basophiles de sorte que la réaction allergique anaphylactique ne se produit pas.
Tolérance immunitaire
La tolérance immunitaire peut être définie comme l'incapacité de l'organisme à développer une réponse immune spécifique contre un antigène donné. Par exemple l'organisme est constitué de protéines et autres substances antigéni-ques; normalement il est tolérant vis-à-vis ses constituants et il ne produit ni anticorps ni lymphocytes T effecteurs contre ses propres antigènes.
La tolérance est causée par l'élimination l'inactivation ou la suppression de lymphocytes B ou T spécifiques résultant d'une exposition particulière à l'antigène. Ainsi dans certaines conditions précises l'exposition à un antigène donné peut empêcher plutôt qu'initier une réponse immune spécifique. La tolérance immunitaire est plus facilement réalisée chez le foetus que chez l'adulte. Au cours du développement foetal l'embryon est presque exclusivement exposé à ses propres antigènes. On croit que durant la vie embryon-naire les antigènes réagissent en paralysant ou en éliminant les clones de lymphocytes B et T particuliers qui auraient pu produire une réponse immune spécifique contre les auto-antigènes. Notons cependant que les lymphocytes B et T continuent de se différencier dans la moelle osseuse et le thymus pendant toute la vie. Le maintien d'une tolérance immunitaire contre ses propres antigènes suppose donc une inhibition active et continue des lympho-cytes B et T qui pourraient induire une réponse immune spécifique contre les auto-antigènes.
On croit que deux mécanismes généraux sont responsables de la tolé-rance: la délétion de clones et la suppression de clones. Dans la délétion clonale, l'exposition à l'antigène élimine ou inactive les clones de lymphocytes B et de lymphocytes T spécifiques à l'antigène. On ne comprend pas encore tout à fait comment ce mécanisme se produit. Cependant dans les conditions appro-priées l'exposition de lymphocytes B à un antigène se traduit apparemment par la disparition ou la transformation des récepteurs capables de fixer l'anti-gène à la surface des lymphocytes B. Ce mécanisme induit une absence de réponse de la part de ces cellules ou une tolérance immunitaire vis-à-vis de cet antigène. Les lymphocytes B immatures sont beaucoup plus sensibles à ce type d'induction que les lymphocytes B matures, et ce fait peut expliquer pourquoi la tolérance est plus facilement induite chez le foetus que chez l'adulte.
Au cours de la suppression clonale, l'exposition particulière à l'antigène active les lymphocytes T suppresseurs qui à leur tour bloquent l'activité des lymphocytes T auxiliaires lesquels participent aussi au déclenchement d'une réponse immune spécifique contre l'antigène. Les lymphocytes T suppresseurs peuvent, d'une part bloquer le développement des lymphocytes T effecteurs responsables des réponses immunes à médiation cellulaire contre l'antigène et peuvent d'autre part, supprimer directement l'activité des lymphocytes B qui pourraient produire une réponse humorale spécifique contre l'antigène. Ainsi dans la suppression clonale les lymphocytes B et les lymphocytes T susceptibles de produire une réponse immune spécifique contre des antigènes particu-liers sont toujours présents mais leur activité est bloquée par les lymphocytes T suppresseurs.
Réponses auto-immunes
Occasionnellement la tolérance immunitaire d'un organisme immunologi-que vis-à-vis de ses propres antigènes s'effondre. Alors apparaissent des réponses iutc-itntriuties. Au cours des réponses auto-immunes l'individu produit des anticorps ou des lymphocytes T effecteurs capables de s'attaquer à ses propres constituants cellulaires. On connaît plusieurs causes capables de déclencher ces phénomènes auto-immuns. Les médicaments les substances chimiques de l'environnement les virus les mutations génétiques peuvent modifier les antigènes normaux et former de nouveaux antigènes à la surface des cellules. L'organisme réagit alors contre ces nouveaux antigènes comme s'il s'agissait d'antigènes étrangers en produisant des anticorps et des lymphocytes T effecteurs pour les éliminer. Des antigènes étrangers ayant une structure chimique apparentée à celle de certains antigènes de l'organisme peuvent également stimuler une production d'anticorps ou de lymphocytes T effecteurs capables de contre-réagir avec les antigènes de l'organisme. Par exemple certains streptocoques possèdent des antigènes qui induisent la formation d'anticorps pouvant contre-réagir avec le tissu cardiaque; des infec-tions streptococciques sévères et répétées produisent parfois un rhumatisme articulaire aigu qui apparaît quelques semaines après l'infection et qui suggère l'existence d'un mécanisme auto-immun. Une autre possibilité serait que les antigènes de l'organisme qui ne sont pas normalement exposés au système immunitaire humoral ou au système immunitaire à médiation cellulaire ne puissent permettre à l'organisme de développer une tolérance immunitaire. Si à la suite d'une lésion tissulaire telle qu'un traumatisme ou une infection ces antigènes sont libérés dans la circulation l'organisme les reconnaît comme antigènes étrangers et réagit en formant des anticorps ou des lymphocytes T effecteurs spécifiquement dirigés contre eux. Par exemple les antigènes de la cornée de l'oeil ne circuleraient pas dans les liquides de l'organisme. Dans le cas d'une lésion de la cornée la libération de ces antigènes peut induire une réponse immune spécifique contre eux qui pourrait causer une opacité de la cornée.
On croit que les lymphocytes T suppresseurs sont très importants pour contrôler les phénomènes d'auto-immunité. Si les lymphocytes T suppresseurs sont associés à la tolérance immunitaire (et particulièrement à la préven-tion de réponses immunes humorales ou à médiation cellulaire contre les auto-antigènes d'un individu) comme on l'a suggéré plus tôt il est concevable qu'une carence de ces lymphocytes puisse contribuer au développement de réponses auto-immunes.
![]()
![]()
![]()
Immunologie: les défenses de l'organisme / virus. | Index |