Défenses de l'organisme contre les infections virales
Besoin de vulgarisation?
 Une info plus large?
Une info plus large?
Aller directement sur la
VIROLOGIE
![]()
I - Moyen de défense de l'organisme contre les infections. Introduction au système immunitaire.
Notre environnement contient une grande diversité de micro-organismes tels que les bactéries, les virus, les champignons ou les parasites. Certains d'entre eux, en se multipliant dans les organismes qui les hébergent peuvent provoquer des désordres graves. Cependant dans la plupart des cas, l'infection s'avère de durée Imitée et l'individu retrouve un état normal. Le système immunitaire, dont l'un des rôles essentiels est de détruire les agents pathogènes et de limiter leur prolifération, est le principal acteur de cette guérison.
Le système immunitaire développe des réactions diverses dont certaines ne sont pas spécifiques de l'agent infectieux. Ce type de réactions qui participent à la formation d'une première barrière de défense contre l'infection constitue l'immunité naturelle (ou immunité non spécifique). Dans un deuxième temps des réactions spécifiques du micro-organisme visé interviennent également pour détruire l'agent infectieux ou les cellules infectées. C'est cette immunité spécifique qui est de surcroît a la base de l'établissement d'une mémoire que conserve l'individu pour éviter toute infection ultérieure par le micro-organisme donné.
Cette immunité qui peut s'installer a vie chez l'individu a la suite de maladies comme les oreillons ou la diphtérie peut également s'acquérir grâce à la vaccination.
I.1 - Moyens de défense non spécifiques
La peau, les sécrétions des glandes sébacées, le mucus, l'acidité gastrique forment diverses barrières physiques et chimiques contre les agents infectieux. Les cellules phagocytaires et les cellules NK ou les cellules nulles qui sécrètent des médiateurs solubles comme l'interféron, font partie de cette immunité cellulaire naturelle.
Les cellules du système immunitaire sont issues de cellules souches pluripotentes suivant deux voies de différenciation: la lignée lymphoïde engendre les lymphocytes B et T qui sont le support de l'immunité spécifique, et d'autres types cellulaires comme les cellules tueuses NK et peut être les cellules nulles qui ne portent aucun de ces deux types de marqueurs.
1.1.1 - Effecteurs de l'activité antivirale
Rôle de l'interleukine 1 dans l'activation des lymphocytes Th (helper)
Principaux mécanismes antiviraux induits par l'interféron (98 k)
I.2 - Moyens de défense spécifiques
Les lymphocytes B et T sont chez les organismes supérieurs le support de l'immunité spécifique. Ils possèdent des récepteurs membranaires capables de reconnaître les antigènes. Les lymphocytes B participent à la formation de l'immunité humorale (réponse anticorps), tandis que les lymphocytes T participent à l'immunité à médiation cellulaire.
- Chez les lymphocytes B, il s'agit d'une immunoglobuline de membrane, classiquement constituée de 2 chaînes légères (L) et de 2 chaînes lourdes (H).
- Les lymphocytes T comportent quant à eux un TCR (T Cell Receptor) constitué de 2 chaînes: alpha bêta ou gamma delta. Le marqueur CD3 est associé au TCR; on parle du complexe TCR/CD3, où le TCR reconnaît spécifiquement l'antigène, tandis que le CD3 active la cellule lorsque le TCR et l'antigène sont associés (ce qui veut dire que l'antigène est reconnu).
1.2.1 - La réponse anticorps (immunité humorale)
Suite à un premier contact avec un antigène, un faible taux d'IgM (immunoglobulines de type M) apparaît après une période de latence, suivi d'une quantité également faible d'IgG. A la suite de cette réponse primaire, les taux d'immunoglobulines décroissent relativement vite. Cependant, en cas de deuxième contact avec l'antigène, la réponse secondaire est caractérisée par une croissance rapide des taux d'anticorps. Ces taux sont plus élevés, plus durables, les anticorps sont plus affins avec une prédominance des IgG sur les IgM.
Un antisérum contre un antigène donné contient une population d'anticorps qui reconnaissent des régions diverses de l'antigène avec des affinités différentes. On parle alors de sérum polyclonal.
Les anticorps monoclonaux sont eux représentés par une population homogène et monospécifique; ils sont obtenus in vitro par la technique des hybridomes. Par leurs caractéristiques, ils sont un outil incomparable pour le diagnostic et éventuellement pour la sérothérapie. Ils permettent également une analyse très fine de la structure antigènique des protéines et des virus.
1.2.1.1 - Structure de base d'une immunoglobuline (138 k, d'après Roitt & coll. Fig.5.3)
1.2.1.2- Origine de la diversité des anticorps
- Les associations VL-VH
Les zones hypervariables (ou CDR) des chaînes lourdes et légères d'une immunoglobuline native sont géographiquement très proches; ce sont elles qui entrent en contact avec l'épitope de l'antigène. La grande variabilité de ces CDR s'associe à un nombre restreint de zones constantes. Par ailleurs, les chaînes VL-VH se trouvent sous une des deux formes possibles kappa ou lambda.
- Les associations VJL-VDJH
Les parties variables s'associent à une courte zone dite de jonction. Celle-ci ne se retrouve pas au niveau de la protéine finale, mais modifie la séquence de la zone variable, en accroissant encore la diversité.
- Les variations somaclonales
Chaque lymphocyte B, bien que stimulés contre un même antigène déterminé, produisent des anticoprs différent sensiblement entre eux. Les anticorps sont dits polyclonaux.
- Les combinaisons de chaînes L/H
Entre deux lymphocytes, les chaînes lourdes (H) peuvent être identiques et les chaînes légères (L) différentes, ou inversement. Ceci entraîne une diversité de combinaisons, d'associations des chaînes qui est à l'origine de la diversité des anticorps.
II -Intéraction Lymphocytes T et Lymphocytes B
II.1 - Reconnaissance des antigènes par les Lymphocytes T.
Les cellules se reconnaissent par le biais de marqueurs, représentés dans la figure ci-dessous. Les molécules LFA 1 des lymphocytes T reconnaissent des marqueurs: ICAM 1, ICAM 2. Il en va de même pour le CD 2, le TRC et le CD 8/4.
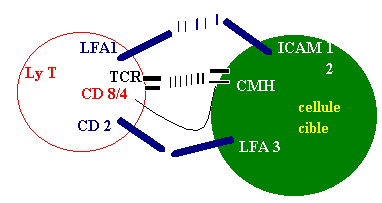
Deux catégories de lymphocytes T se trouvent impliquées dans
ce type d'intéraction: les Ly T4 et les Ly T8. Les premiers ne
possèdent qu'un CD 4, dont le rôle est de reconnaître
une classe de molécules du CMH:, celles de classe II (voir schéma).
Avec la participation de l'interleukkine 1 les Ly T4 sont activés
en Ly T helpers producteurs d'interleukine 2. Cette dernière active
et 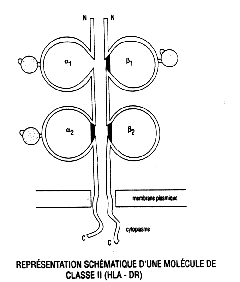 environnement contient une grande diversité de micro-organismes tels
que les bactéries, les virus, les champignons ou les parasites. Certains
d'entre eux, en se multipliant dans les organismes qui les hébergent
peuvent provoquer des désordres graves. Cependant dans la plupart
des cas, l'infection s'avère de durée Imitée et l'individu
retrouve un état normal. Le système immunitaire, dont l'un
des rôles essentiels est de détruire les agents pathogènes
et de limiter leur prolifération, est le principal acteur de cette
guérison.
environnement contient une grande diversité de micro-organismes tels
que les bactéries, les virus, les champignons ou les parasites. Certains
d'entre eux, en se multipliant dans les organismes qui les hébergent
peuvent provoquer des désordres graves. Cependant dans la plupart
des cas, l'infection s'avère de durée Imitée et l'individu
retrouve un état normal. Le système immunitaire, dont l'un
des rôles essentiels est de détruire les agents pathogènes
et de limiter leur prolifération, est le principal acteur de cette
guérison.
II.2 - Actvation des cellules T par les Cellules Présentatrices de l'Antigène (CPA)
La figure ci-dessous illustre comment les cellules du système immunitaire coopèrent entre elles par l'intermédiaire des interleukines. Ces molécules ne sont pas spécifiques de l'antigène et agissent à de très faibles concentration de l'ordre du picomolaire.
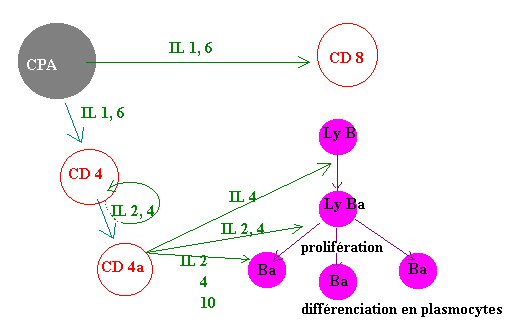
L'activité des interleukines (IL) - et des cytokines en général - est pléïotropique, c'est à dire qu'ils ont un très large spectre d'action selon la cellule cible: un ostéoclaste n'engendrera pas la même réponse qu'une cellule épithéliale par exemple. Les cytokines sont stimulées en réponse à une stimulation (antigénique) et les cellules cibles de ces cytokines n'y sont sensibles (autrement dit ne synthétisent le récepteur membranaire) que si elles ont été activées au préalable (par une autre cytokine par exemple).
Les cytokines permettent l'activation, l'inhibition ou la prolifération cellulaire; ces réponses sont fonction de la spécificité de la cellule cible.
Lorssque le système immunitaire est dépassé, il recrute (par l'intermédiaire des cytokines) des cellules non spécifiques tels que les macrophages et les polynucléaires.
![]()
![]()